
DNAポリメラーゼの定義
DNAポリメラーゼは、デオキシリボヌクレオチドからのDNA分子の合成を触媒する酵素です。 DNAポリメラーゼは、世代から世代へと遺伝情報を娘細胞に渡すことを可能にするDNA複製において重要な役割を果たします。
Christina Swords、Ph.D。が編集
DNAポリメラーゼの生化学的側面
ポリメラーゼ活性
ポリメラーゼは、個々の分子(モノマー)の化学的結合が鎖(ポリマー)を形成することを可能にします。 DNAポリメラーゼの場合、形成されるポリマーはデオキシリボ核酸(DNA)です。 モノマーはデオキシリボヌクレオチド、より正確にはデオキシヌクレオシド三リン酸(dNTP)です。 DNA依存性DNAポリメラーゼは、常に既存のDNA一本鎖をテンプレートとして使用し、そのヌクレオチド配列がテンプレートによって決定される新しい相補鎖を合成します。 DNA配列のこの保存は、DNAポリメラーゼがDNAにコードされている遺伝情報をコピーする能力にとって決定的です。 テンプレートの正しいコピーは、水素結合によって媒介される、組み込まれたヌクレオチド塩基とDNAテンプレートの塩基との相補的な塩基対形成によって達成されます。 新しいDNA鎖の合成は、5 ‘から3’の末端まで行われます。 化学的には、dNTPのαリン酸に対するDNA鎖の末端3’-ヒドロキシ基の求核攻撃が起こり、ピロリン酸が放出されます。 このステップは、ポリメラーゼによって触媒されます。
RNAポリメラーゼ(アミノ酸からタンパク質を合成するために使用されるRNAを生成する)とは対照的に、DNAポリメラーゼでの相補的DNA鎖の合成は、ポリメラーゼが遊離の3′-ヒドロキシ末端を利用できる場合にのみ行うことができます。 次に、最初のヌクレオチドがこの末端に結合します。 ポリメラーゼ連鎖反応(PCR)では、長さが約15〜20ヌクレオチドのDNA一本鎖(プライマー)が反応の開始点として使用されます。 酵素は通常、補因子としてマグネシウムイオンを必要とします。
ジエステル結合の形成の触媒作用は、RNAポリメラーゼの対応する反応と機能的に類似しています。 すでに合成されたセクションの最後のヌクレオチドと追加されるヌクレオチドは、それぞれポリメラーゼドメインの触媒中心にある2つのマグネシウムイオンの1つに配位しています。 追加されるヌクレオチドの最初のリン酸基は、両方のマグネシウムイオンに配位しています。 空間的位置により、先行するヌクレオチドのヒドロキシ基が、付加されるヌクレオチドのリン酸基を攻撃することが可能になる。 その過程で、ピロリン酸残基が分離されます。
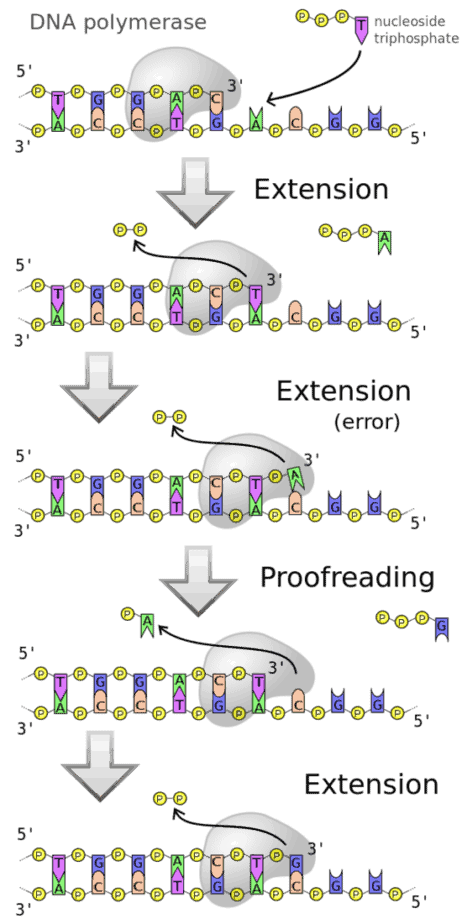
エキソヌクレアーゼ活性
多くのポリメラーゼは他の酵素機能も持っています。 低濃度のdNTPの存在下では、ヌクレオチド除去のための3 ‘→5’エキソヌクレアーゼ活性が優勢です。 一部のポリメラーゼは、5 ‘→3’エキソヌクレアーゼ活性も持っています。 DNAテンプレートの読み取り時にエラーが発生しないようにするために、この校正機能があります。つまり、不適切なヌクレオチドの挿入を検出し、エキソヌクレアーゼ活性によってDNAから除去することができます。 これにより、新しい鎖が形成されている間に、テンプレート鎖とすでにペアになっている既存のDNAまたはRNA鎖の分解が可能になります。 これにより、古いストランドが新しいストランドに交換されます。 このエキソヌクレアーゼ活性は、ニックトランスレーション法によって利用されます。
さまざまなDNAポリメラーゼ
大腸菌などの細菌には、3つの異なるDNA依存性DNAポリメラーゼがあります。 それらの1つであるDNAポリメラーゼI(Pol I)は、1955年にArthur Kornbergによって分離され、これまでに発見された最初のポリメラーゼでした。 ただし、これは約20の合成ステップしか触媒しないため、大腸菌での複製に最も重要なポリメラーゼではありません(つまり、処理能力が低いだけです)。 ただし、5 ‘→3’エキソヌクレアーゼ活性のため、複製中のプライマー分解の原因となります。 大腸菌の他の2つのDNAポリメラーゼであるDNAポリメラーゼIIとDNAポリメラーゼIIIは、DNAポリメラーゼIの発見からわずか15年後に分離されました。その後、ポリメラーゼI遺伝子に欠陥があるE.coli変異体が複製。 ただし、これらの変異体は特に紫外線やアルキル化物質の影響を受けやすいため、DNAポリメラーゼIが主に修復作業を行うと考えられています。 大腸菌で実際の複製を行うポリメラーゼIIIは、合計7つのサブユニットで構成されており、細菌細胞あたりのコピー数はごくわずかです。
ヒトDNAポリメラーゼを含む真核生物のDNAポリメラーゼは、次のファミリーに分類されます。
- ファミリーA:DNAポリメラーゼγ、θ、およびν
- ファミリーB:DNAポリメラーゼα、δ、εおよびζ
- ファミリーX:DNAポリメラーゼβ、λ、σおよびμ
- ファミリーY:DNAポリメラーゼη、ιおよびκ
ポリメラーゼγはミトコンドリアでのみ発生します。
哺乳類では、α、β、γ、δ、εの5種類しか発生しません。 複製の決定要因であるポリメラーゼδおよびεは、高い処理能力と校正機能を特徴としていると考えられます。 対照的に、ポリメラーゼαおよびβは、低い処理能力のみを示し、校正機能を示さない。
さらに、RNAをテンプレートとして使用し、それにdNTPを付加するRNA依存性DNAポリメラーゼがあります。 これらは逆転写酵素と呼ばれ、テロメラーゼも含まれます。 唯一知られている独立したDNAポリメラーゼは、ターミナルデオキシリボヌクレオチジルトランスフェラーゼです。
古細菌には、PCRにも使用される温度安定型があります。
生物学的意義
DNAポリメラーゼは、DNA複製にとって最も重要です。 それらは、DNAの形で遺伝子情報の忠実なコピーを可能にし、したがって、生物の繁殖と生殖における決定的なステップを可能にします。 酵素はまた、DNA修復に関連するプロセスにおいて重要な役割を果たします。
生物工学的意義
研究室では、DNAポリメラーゼは、ポリメラーゼ連鎖反応および関連する方法(RT-PCR、qPCRなど)、ニックトランスレーション、ランダムプライミング、およびDNAシーケンシングによく使用されます。 多数の異なる耐熱性タイプ(例:TaqポリメラーゼThermus aquaticus )が使用され、その一部はタンパク質工学によって変更されています。 高温安定性に加えて、Pfuポリメラーゼなどの古風な起源の熱安定性DNAポリメラーゼは、PCRが生成されたDNAの変化を引き起こしてはならないため、証拠の読み取りを提供します。 さらに、φ29DNAポリメラーゼなどの鎖置換DNAポリメラーゼは、室温での等温DNA増幅のさまざまな方法で使用されます。 今日使用されているDNAポリメラーゼの前駆体は、T4DNAポリメラーゼでした。
このブログ投稿は気に入りましたか? 発見できるDNA、遺伝学、シーケンシングに関するその他の投稿はこちら!
次の関連ブログ投稿に興味があるかもしれません。