Введение в ДНК
Дезоксирибонуклеиновая кислота — это нуклеиновая кислота, которая представляет собой полинуклеотид, состоящий из цепочки из многих нуклеотидов. Биомолекула, расположенная в хромосомах, является носителем генетической информации во всех живых организмах и многих вирусах (ДНК-вирусы, параретровирусы), т.е. материальной основой генов.
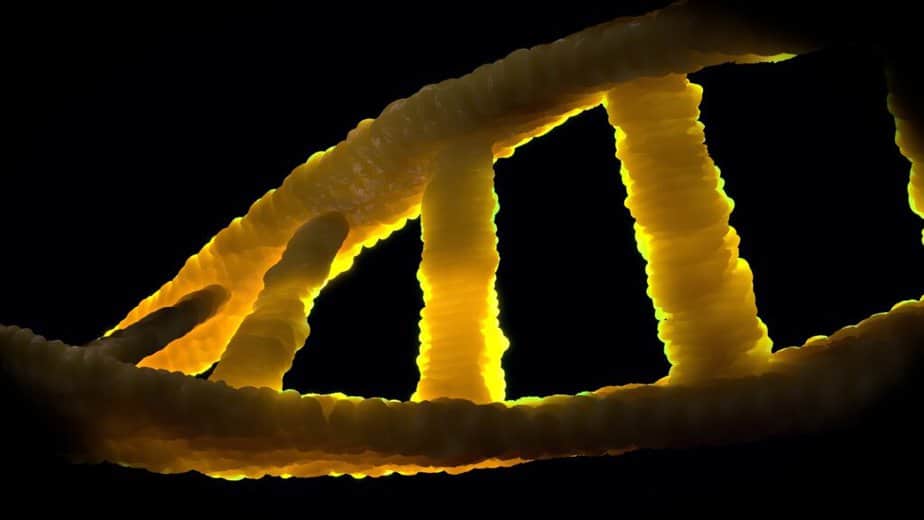
В нормальном состоянии ДНК имеет форму двойной спирали. Его строительные блоки представляют собой четыре разных нуклеотида, каждый из которых состоит из фосфатного остатка, сахарной дезоксирибозы и одного из четырех органических оснований (аденина, тимина, гуанина и цитозина, часто обозначаемых аббревиатурой A, T, G и C).
Гены в ДНК содержат информацию о производстве рибонуклеиновых кислот. В генах, кодирующих белок, это важная группа РНК, мРНК. Он, в свою очередь, содержит информацию для построения белков, которые необходимы для биологического развития живого существа и метаболизма в клетке. Последовательность оснований здесь определяет последовательность аминокислот соответствующего белка: генетический код кодирует конкретную аминокислоту с тремя соседними основаниями каждая.
В клетках эукариот, к которым также относятся растения, животные и грибы, большая часть ДНК в ядре клетки (латинское ядро, следовательно, ядерная ДНК или яДНК) организована в виде хромосом. Небольшая часть находится в митохондриях, «электростанциях» клеток, и соответственно называется митохондриальной ДНК (мтДНК). У растений и водорослей также есть ДНК в фотосинтетических органеллах, хлоропластах или пластидах (хпДНК). У бактерий и архей — прокариот, не имеющих клеточного ядра, — ДНК находится в цитоплазме. Некоторые вирусы, так называемые РНК-вирусы, хранят свою генетическую информацию в РНК, а не в ДНК.

Под редакцией Кристины Сордс, доктора философии.
Структура и организация ДНК
Строительные блоки
Дезоксирибонуклеиновая кислота — это длинноцепочечная молекула (полимер), состоящая из множества строительных блоков, называемых дезоксирибонуклеотидами или для краткости нуклеотидами. Каждый нуклеотид состоит из трех компонентов: фосфорной кислоты или фосфата, дезоксирибозы сахара и гетероциклического азотистого основания или основания. Субъединицы дезоксирибозы и фосфорной кислоты одинаковы для каждого нуклеотида. Они составляют основу молекулы. Единицы основания и сахара (без фосфата) называются нуклеозидами.
Остатки фосфата гидрофильны из-за их отрицательного заряда; они придают ДНК в водном растворе общий отрицательный заряд. Поскольку эта отрицательно заряженная ДНК, растворенная в воде, не может больше испускать протоны, это, строго говоря, не (уже не) кислота. Термин дезоксирибонуклеиновая кислота относится к незаряженному состоянию, в котором протоны присоединены к фосфатным остаткам.
Основание может представлять собой пурин, а именно аденин (A) или гуанин (G), или пиримидин, а именно тимин (T) или цитозин (C). Поскольку четыре разных нуклеотида различаются только своим основанием, сокращения A, G, T и C также используются для соответствующих нуклеотидов.
Пять атомов углерода дезоксирибозы пронумерованы от 1 до 5. Основа привязана к концу этого сахара. Остаток фосфата прикреплен к 5 ‘концу. Строго говоря, дезоксирибоза — это 2-дезоксирибоза; Название происходит от того факта, что по сравнению с молекулой рибозы спиртовая гидроксигруппа (группа ОН) отсутствует в положении 2 ‘(то есть заменена атомом водорода).
В 3′-положении находится группа ОН, которая связывает дезоксирибозу с 5′-атомом углерода сахара следующего нуклеотида через так называемую фосфодиэфирную связь. Таким образом, каждая так называемая одинарная нить имеет два разных конца: конец 5 ‘и конец 3’. ДНК-полимеразы, которые синтезируют нити ДНК в живом мире, могут присоединять новые нуклеотиды только к ОН-группе на 3′-конце, но не на 5′-конце. Таким образом, одиночная нить всегда вырастает с 5 футов до 3 футов. Нуклеозидтрифосфат (с тремя фосфатными остатками) поставляется как новый строительный блок, от которого отделяются два фосфата в форме пирофосфата. Оставшийся фосфатный остаток каждого вновь добавленного нуклеотида соединяется с группой ОН на 3′-конце последнего нуклеотида, присутствующего в цепи, путем отщепления воды. Последовательность оснований в цепи кодирует генетическую информацию.
Двойная спираль
ДНК обычно представляет собой спиральную двойную спираль в конформации, называемой B-ДНК. Две из описанных выше одинарных нитей прикреплены друг к другу в противоположных направлениях: на каждом конце двойной спирали одна из двух одинарных нитей имеет свой 3 ‘конец, а другая — 5’ конец. Из-за наложения двух специфических оснований в середине двойной спирали всегда напротив друг друга, они «спарены». Двойная спираль в основном стабилизируется за счет взаимодействия между последовательными основаниями одной и той же цепи (а не, как часто утверждают, водородными связями между нитями).
Аденин и тимин всегда спарены, образуя две водородные связи, или цитозин с гуанином, которые связаны тремя водородными связями. Мостик образуется между положениями молекулы 1 = 1 и 6 = 6, а в случае пар гуанин-цитозин дополнительно между 2 = 2. Поскольку одни и те же основания всегда являются парными, последовательность оснований в одной цепи может использоваться для получения оснований другой цепи, последовательности являются комплементарными. Водородные связи почти исключительно ответственны за специфичность спаривания, но не за стабильность двойной спирали.
Поскольку пурин всегда сочетается с пиримидином, расстояние между нитями везде одинаково, образуется правильная структура. Вся спираль имеет диаметр около 2 нм и закручивается на 0,34 нм с каждой молекулой сахара.
Плоскости молекул сахара расположены под углом 36 ° друг к другу, поэтому полное вращение достигается после 10 оснований (360 °) и 3,4 нм. Молекулы ДНК могут стать очень большими. Например, самая большая хромосома человека содержит 247 миллионов пар оснований.
Когда две отдельные пряди скручиваются вместе, остаются боковые зазоры, так что основания здесь находятся прямо на поверхности. Две из этих канавок наматываются вокруг двойной спирали (см. Иллюстрации и анимацию в начале статьи). «Большая борозда» имеет ширину 2,2 нм, «малая борозда» — всего 1,2 нм.
Соответственно, основания в большой борозде более доступны. Белки, которые связываются с ДНК специфическим для последовательности образом, такие как факторы транскрипции, поэтому обычно связываются с большой бороздкой.
Некоторые красители ДНК, например DAPI, также связываются с бороздой.
Кумулятивная энергия связи между двумя одиночными нитями удерживает их вместе. Ковалентных связей здесь нет, а это значит, что двойная спираль ДНК состоит не из одной молекулы, а из двух. Это позволяет временно разделить две нити в биологических процессах.
В дополнение к только что описанной B-ДНК существует также A-ДНК и левосторонняя, так называемая Z-ДНК, которая была впервые изучена в 1979 году Александром Ричем и его коллегами из Массачусетского технологического института. Это особенно характерно для участков с высоким содержанием GC. Лишь в 2005 г. появилась информация о кристаллической структуре, которая показывает Z-ДНК непосредственно в соединении с B-ДНК и, таким образом, обеспечивает доказательства биологической активности Z-ДНК. В следующей таблице и на рисунке рядом показаны различия между тремя формами в прямом сравнении.
Базовые стопки не совсем параллельны друг другу, как книги, а образуют клинья, которые наклоняют спираль в ту или иную сторону. Самый большой клин образован аденозинами в паре с тимидинами другой цепи. Следовательно, серия пар AT образует дугу в спирали. Когда такие серии следуют друг за другом через короткие промежутки времени, молекула ДНК принимает изогнутую или изогнутую структуру, которая является стабильной. Это также называется дифракцией, индуцированной последовательностью, поскольку дифракция также может быть вызвана белками (так называемая дифракция, индуцированная белками). Дифракция, вызванная последовательностью, часто обнаруживается в важных местах генома.
Хроматин и хромосомы
Хромосомы человека в поздней метафазе деления ядра митотической клетки: каждая хромосома показывает две хроматиды, которые разделены в анафазе и разделены на два ядра клетки.
В эукариотической клетке ДНК организована в виде нитей хроматина, называемых хромосомами, которые расположены в ядре клетки. Одиночная хромосома содержит длинную непрерывную двойную цепь ДНК (в хроматиде) от анафазы до начала S-фазы. В конце S-фазы хромосома состоит из двух идентичных цепей ДНК (в двух хроматидах).
Поскольку такая нить ДНК может иметь длину несколько сантиметров, а ядро клетки — всего несколько микрометров в диаметре, ДНК необходимо дополнительно сжать или «упаковать». У эукариот это делается с помощью так называемых белков хроматина, из которых особенно примечательны основные гистоны. Они образуют нуклеосомы, вокруг которых на самом низком уровне упаковки обернута ДНК. Во время деления ядра (митоза) каждая хромосома конденсируется до максимально компактной формы. Это позволяет их особенно хорошо идентифицировать под световым микроскопом во время метафазы.

Бактериальная и вирусная ДНК
В прокариотических клетках большая часть двухцепочечной ДНК в случаях, задокументированных до сих пор, представлена не в виде линейных цепей с началом и концом каждая, а в виде кольцевых молекул — каждая молекула (т.е. каждая цепь ДНК) замыкает круг со своими 3 ‘и 5’ конец. Эти две кольцевые замкнутые молекулы ДНК называются бактериальной хромосомой или плазмидой, в зависимости от длины последовательности. У бактерий они также не расположены в ядре клетки, а свободно присутствуют в плазме. Прокариотическая ДНК сворачивается в простые «суперспирали» с помощью ферментов (например, топоизомеразы и гиразы), которые напоминают кольцевой телефонный шнур. По-прежнему вращая спирали вокруг себя, пространство, необходимое для генетической информации, уменьшается. В бактериях топоизомеразы гарантируют, что скрученная двойная цепь оторвана в желаемом месте, постоянно разрезая и воссоединяя ДНК (предварительное условие для транскрипции ДНК и репликации ДНК). В зависимости от типа вирусы содержат в качестве генетической информации ДНК или РНК. И в ДНК, и в РНК-вирусах нуклеиновая кислота защищена белковой оболочкой.
Химические и физические свойства структуры двойной спирали ДНК
При нейтральном pH ДНК представляет собой отрицательно заряженную молекулу, причем отрицательные заряды находятся на фосфатных группах в основной цепи цепей. Хотя две из трех кислотных ОН-групп фосфатов этерифицированы соответствующими соседними дезоксирибозами, третья все еще присутствует и высвобождает протон при нейтральном pH, что вызывает отрицательный заряд. Это свойство используется в электрофорезе в агарозном геле для разделения различных цепей ДНК в зависимости от их длины. Некоторые физические свойства, такие как свободная энергия и температура плавления ДНК, напрямую связаны с содержанием GC, т.е. они зависят от последовательности.
Взаимодействия стека ДНК
Два основных фактора несут ответственность за стабильность двойной спирали: спаривание оснований между комплементарными основаниями и взаимодействия стэкинга между последовательными основаниями.
Вопреки первоначальному предположению, выигрыш в энергии за счет водородных связей незначителен, поскольку основания могут образовывать такие же хорошие водородные связи с окружающей водой. Водородные связи пары оснований GC вносят лишь минимальный вклад в стабильность двойной спирали, тогда как связи пары оснований AT даже имеют дестабилизирующий эффект. С другой стороны, стэковые взаимодействия имеют эффект только в двойной спирали между последовательными парами оснований: между ароматическими кольцевыми системами гетероциклических оснований образуется дипольное дипольное взаимодействие, которое является энергетически выгодным. Таким образом, образование первой пары оснований весьма неблагоприятно из-за низкого прироста и потерь энергии, но удлинение (удлинение) спирали является энергетически выгодным, поскольку стэкинг пар оснований происходит при выигрыше энергии.
Однако стэкинг-взаимодействия зависят от последовательности и энергетически наиболее благоприятны для укладываемого GC-GC, менее благоприятны для укладываемого AT-AT. Различия в стэкинг-взаимодействиях в основном объясняют, почему GC-богатые участки ДНК термодинамически более стабильны, чем AT-богатые участки, в то время как водородные связи играют второстепенную роль.
Точка плавления ДНК
Точка плавления ДНК — это температура, при которой преодолеваются силы связывания между двумя одиночными цепями и они отделяются друг от друга. Это также называется денатурацией.
Пока ДНК денатурируется при кооперативном переходе (который происходит в узком температурном диапазоне), точкой плавления является температура, при которой половина двойных цепей денатурируется в одиночные цепи. Из этого определения выводятся правильные термины «средняя точка температуры перехода» Tm.
Температура плавления зависит от соответствующей последовательности оснований в спирали. Он увеличивается, если в спирали больше пар оснований GC, поскольку они энтропийно более выгодны, чем пары оснований AT. Это связано не столько с разным количеством водородных связей, образованных двумя парами, сколько с разными взаимодействиями стэкинга. Энергия суммирования двух пар оснований намного меньше, если одна из двух пар является парой оснований AT. С другой стороны, стеки GC энергетически более выгодны и сильнее стабилизируют двойную спираль. Отношение пар оснований GC к общему количеству всех пар оснований определяется содержанием GC.
Поскольку одноцепочечная ДНК поглощает УФ-свет примерно на 40 процентов сильнее, чем двухцепочечная ДНК, температуру перехода можно легко определить с помощью фотометра.
Если температура раствора упадет ниже Tm, отдельные нити могут снова соединиться друг с другом. Этот процесс называется ренатурацией или гибридизацией. Взаимодействие де- и ренатурации используется во многих биотехнологических процессах, например, в полимеразной цепной реакции (ПЦР), саузерн-блоттинге и гибридизации in situ.
Вам понравился этот пост в блоге? Ты можешь найти больше сообщений о ДНК, генетике и секвенировании здесь !
Возможно, вас заинтересуют следующие статьи блога по теме:
Перед отъездом ознакомьтесь с нашим секвенированием всего генома!